






























 Arguments
Arguments


















Coral Reef Baselines
Posted on 12 February 2011 by John Bruno
The pristine or natural state of a population or community is called the baseline in conservation biology. A baseline serves as a guide for setting conservation and restoration targets. Unfortunately, scientists rarely have reliable information on baselines, because in most cases quantitative data are not collected until long after the resource has been modified.
A new paper in Coral Reefs “Assessing loss of coral cover on Australia’s Great Barrier Reef over two decades, with implications for longer-term trends” (Sweatman et al. 2011) tries to get at what the baseline is for the Great Barrier Reef (GBR) using the results of ecological surveys performed by the Australian Institute of Marine Science (AIMS). Sweatman et al argue that the AIMS surveys, which began in 1986, are the most reliable evidence we have and that other evidence should be ignored. However, many other scientists surveyed reefs on the GBR (for various reasons) decades before AIMS began it’s monitoring program.
Three papers have collated that data and combined it with the AIMS survey data to estimate how the GBR has changed over the last 4-5 decades. The first such paper (published in Nature by Bellwood et al 2004) included this graphic of long-term change in coral cover (the percentage of the sea floor covered by living corals – because corals facilitate so many reef inhabitants, living coral cover is a key measure of reef habitat quality and quantity, analogous to the coverage of trees as a measure of tropical forest loss):
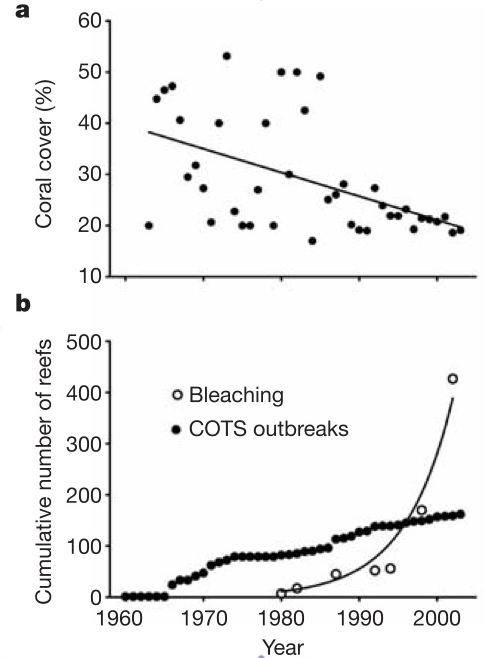
Figure 1 (from Bellwood et al 2004). Degradation of coral reefs. a, Results of a meta-analysis of the literature, showing a decline in coral cover on the Great Barrier Reef. Each point represents the mean cover of up to 241 reefs sampled in each year. b, The recorded number of reefs on the Great Barrier Reef, Australia, substantially damaged over the past 40 yr by outbreaks of crown-of-thorns starfish (COTS) and episodes of coral bleaching.
The second study (Bruno and Selig 2007) is a meta-analyses of coral reef survey data from 2667 reefs across the Indo-Pacific performed between 1968 and 2004 (Fig. 2).

Figure 2 (from Bruno and Selig 2007). Coral cover in ten Indo-Pacific subregions in each of three periods. Plotted values are means +/- 1 SE and values above each bar are the subregional sample sizes. *=no data available
The third paper, Pandolfi et al 2003, used a variety of historical and paleontological data sources in an attempt to reconstruct the longer-term, including pre-human, history of the GBR and other reefs around the world (Fig. 3).

Figure 3 (from Pandolfi 2003). Time trajectories for reef regions over seven cultural periods.
All three studies essentially concluded that GBR coral cover and overall ecosystem health began to decline decades ago, despite the fact that the GBR is currently in better shape than many of the world’s reefs.
Sweatman et al dismiss pre-1986 data from all three studies, arguing that only AIMS survey data are suitable, and thus conclude that the GBR has changed little if at all due to human influences: "We argue that the GBR is currently less degraded from its natural, resilient state than some published reports have asserted".
Global climate change skeptics have frequently use a very similar approach: they rationalize cherry picking a favored data set and time interval in an attempt to show land and ocean temperatures haven’t increased, that sea ice hasn’t declined, etc.
Abstract (from Sweatman et al 2011): While coral reefs in many parts of the world are in decline as a direct consequence of human pressures, Australia’s Great Barrier Reef (GBR) is unusual in that direct human pressures are low and the entire system of 2,900 reefs has been managed as a marine park since the 1980s. In spite of these advantages, standard annual surveys of a large number of reefs showed that from 1986 to 2004, average live coral cover across the GBR declined from 28 to 22%. This overall decline was mainly due to large losses in six (21%) of 29 subregions. Declines in live coral cover on reefs in two inshore subregions coincided with thermal bleaching in 1998, while declines in four mid-self subregions were due to outbreaks of predatory starfish. Otherwise, living coral cover increased in one subregion (3%) and 22 subregions (76%) showed no substantial change. Reefs in the great majority of subregions showed cycles of decline and recovery over the survey period, but with little synchrony among subregions. Two previous studies examined long-term changes in live coral cover on GBR reefs using meta-analyses including historical data from before the mid-1980s. Both found greater rates of loss of coral and recorded a marked decrease in living coral cover on the GBR in 1986, coinciding exactly with thestart of large-scale monitoring. We argue that much of the apparent long-term decrease results from combining data from selective, sparse, small-scale studies before 1986 with data from both small-scale studies and large-scale monitoring surveys after that date. The GBR has clearly been changed by human activities and live coral cover has declined overall, but losses of coral in the past 40–50 years have probably been overestimated.

For several reasons listed below, I think the main conclusion of Sweatman et al is unsupported once you consider the scientific record as a whole.
1) There is much more pre-1986 data available than Sweatman et al suggest
Our study alone (Bruno and Selig) includes data from 154 surveys of reefs across the Indo Pacific performed between just 1980 and 1982. We found that mean coral cover was 42.5% (95% CI, 39.3 and 45.6). Our analysis includes 104 GBR surveys performed between 1968 and 1983, all from published literature (see Fig. 2).
2) Baseline data from other regions indicates that historically GBR coral cover was higher than it is today (or was in 1986)
In the Caribbean a small number (a few dozen) of reliable quantitative surveys from before the early 1980s suggest the regional mean for coral cover was 30-40% (Gardner et al 2003, Schutte et al 2010: see Figs. 5 and 6 below). Because countless well trained reef scientists were working throughout the region and observed the state of reefs over the last 100 years, most Caribbean reef scientists think that historically, the regional mean of Caribbean coral cover was higher than this; probably closer to 50% (or greater). Most Caribbean reefs of this era were dominated by Acropora spp., as can be seen in the photo below from 1974:

Sweatman et al are effectively arguing that the GBR has naturally lower coral cover (averaging a mere 28%) than the Caribbean, which lacks plating species; even in dense thickets of branching Acroporids (as seen above), Caribbean coral cover rarely exceeds 70% whereas on the GBR, cover can easily reach 100% (see the photo below).

We also have substantially more older survey data for several non-GBR Pacific regions than are available for the GBR. For example, Gomez et al surveyed more than 600 sites in the Philippines in 1981 (Gomez et al 1981). Their work clearly shows a coral baseline far higher than today in the Philippines, the GBR or anywhere else in the world. Also see Fig. 4 below from Bruno and Selig (2010) that suggests the values reported in Gomez et al are representative of other regions at the time. Also note the striking difference in the distribution of coral cover values among reefs between the early 1980s and for more recent surveys. I don't think it is logical to assume that the GBR hasn't followed these global trends and that it has a much lower coral cover baseline than the rest of the world.

Figure 4. (From Bruno and Selig 2007) Histograms illustrating percent coral cover in the Indo-Pacific and selected subregions during different periods.
3) The GBR was already disturbed and changing by the time the AIMS monitoring program began
AIMS began surveying a few dozen reefs along the GBR in 1986 after it became obvious that the reef ecosystem was changing due to over-fishing, sediment pollution from coastal development, ocean warming and predator outbreaks. Sweatman et al argue the values recorded by the early AIMS LTMP surveys of the late-1980s, when cover was roughly 30%, are representative of the pre-human, historical baseline. I see no reason to assume this given the well-documented anthropogenic disturbances that were already affecting the reef by then (see Fig 1).
4) The early GBR survey data are sound
Sweatman et al: Another concern is that the objectives of many early reef studies predisposed them to select areas of high coral cover...This change in research emphasis is likely to be reflected in the choice of study sites, with early studies selecting sites for their diverse communities and high coral cover where the study organisms are abundant and large samples can be found in a small area.I initially thought this too, but once I got into the literature, I was reminded that the late 60s and early 70s were the heyday of disturbance ecology and most reef ecologists were community ecologists, and like their peers working in other systems, focused almost entirely on the effects of large disturbances on reef communities, e.g., Endean and Stablum 1973, Endean 1977, Connell 1978, Done 1992, Done et al. 1991, etc. Thus, if anything, I think most the early bias is in the other direction. Regardless, I agree site selection biases, both then and now, complicate long-term trend interpretation.
Sweatman et al: While the AIMS long-term data show a decline in average coral cover on GBR reefs from 28.1 to 21.7% between 1986 and 2004, two studies (Bellwood et al. 2004; Bruno and Selig 2007) based on unweighted meta-analyses have suggested that average coral cover on GBR reefs was considerably higher in the 1960s and 1970s than in the 1980s. Bellwood et al. (2004) presented a plot of mean coral cover on the GBR indicating that cover halved from ~40% in the early 1960s to ~20% in 2000, almost three times the rate of decline in the AIMS long-term monitoring data. Bruno and Selig (2007) used information from 2,667 sites to assess change across the Indo-Pacific, including the GBR as one subregion. Based on published studies including AIMS monitoring data, they found that mean coral cover on the GBR declined by ~25% (in relative terms) from the period 1968–1983 to 1984–1996 and then was relatively stable until the end of their study period in 2004. We argue that this difference is substantially due to a change in the scale of surveys and in survey methods. The most compelling evidence for this is that, in the data sets of both studies, the annual estimates of the mean for coral cover on the GBR drop abruptly in 1986, the first year of large-scale moni- toring on the GBR, and then vary rather little around the new level in subsequent years. The apparent abrupt drop in average coral cover on the GBR in 1986 is most probably due to the inclusion of AIMS monitoring data with cover estimates from small selected patches of reef from small-scale studies...Like Sweatmean et al, Bruno and Selig and Bellwood et al found that at a regional scale, average coral cover on the GBR has changed little since 1984 (see Figs. 1 and 2 above). A similar pattern has been documented for the Caribbean, where substantial coral loss in the early 1980s changed to relative regional stasis since (Figs. 5A and C), i.e., a very similar pattern has been found in other regions (regardless of the scale of surveys and survey methods - also see Bruno and Selig for other Indo-Pacific examples) in which AIMS has never surveyed.

Figure 5 (From Schutte et al 2010). Annual cover values (±1 SE, closed circles, left y-axis) and site sample sizes (open circles, right y-axis) for (A) mean coral cover for all sites in the Caribbean basin (n = 1962; star: 1980, the year in which Hurricane Allen struck and white band disease outbreaks began); (B) mean macroalgal cover for all sites for which data were available (n = 875; star: 1983, the year in which the Diadema antillarum die-off began); (C) mean coral cover for all sites in the greater Caribbean except those in the Florida Keys (FLK; n = 1515); and (D) mean coral cover for all sites in the FLK subregion (n = 447)
The AIMS survey data presented in Sweatman et al used the "manta tow" technique; snorkelers are towed behind a boat, over a reef and visually estimate coral cover within 10m wide swaths to categories such as 0-5%, >5-10%, 10-20%, 20-30%, etc. I think the manta technique can be a valid tool for estimating crude spatio-temporal trends in coral cover. I even used the AIMS manta data in our Bruno and Selig paper, although I haven't used it since (having spoken with several former AIMS technicians about the technique and it's accuracy and precision). In short, Sweatman et al argue that their manta tow data is so superior to data from other sources, that only the AIMS manta data should be used to asses long-term trends in GBR state. I disagree; not to knock the AIMS manta data, but it is a visual quasi-quantitative estimate collected by technicians as compared to the pre-1986 GBR data which was all collected by renouned PhD scientists, e.g., Endean and Stablum, Connell, Done, etc, that spent their lives studying the GBR.
Furthermore, these pre-1986 GBR surveys were performed using wholly quantitative techniques such as underwater photography to estimate coral cover. I just don't buy the argument that the AIMS-intern-manta tow data are superior to quantitative surveys by trained PhD scientists.

Figure 6 (From Gardner et al 2003). Absolute percent coral cover from 1977 to 2001. Annual coral cover estimates (black triangles) are weighted means with 95% bootstrap confidence intervals. Also shown are unweighted mean coral cover estimates for each year (black circles), the unweighted mean coral cover with the Florida Keys Coral Monitoring Project (1996–2001) omitted (X), and the sample size (number of studies) for each year (white circles, right y axis).
Other skeptic soundbites
Sweatman et al make a lot of noise about variation among reefs and subregions in coral cover trajectories and states. Basically, there is a high degree of spatio-temporal asynchrony in the system, as we discussed in Bruno and Selig and illustrted with this graphic:

Figure 7 (from Bruno and Selig 2007). Illustrative examples of asynchrony of coral cover among 25 randomly selected monitored reefs on the GBR (a) and in Indonesia (b).
This isn't surprising or atypical and it doesn't mean a long-term trend isn't present or detectable, as Sweatman et al suggest. Climate change deniers use this argument frequently, suggesting that natural short term variation makes long-term, anthropogenically forced trends, unlikely or undetectable. This is in a sense what Sweatman et al are arguing as well. But the fact that there is great unforced spatiotemporal variation, i.e, noise or weather, does not mean that longer-term, human-induced change isn't also happening. (Although this is indeed a big issue for many fields of global change science: how to detect slow, long-term trends in a sea of shorter term noise.)Sweatman et al: A reef system that is stable in the long term will still show cycles of disturbance and recovery at a subregional scaleTrue, but so will a reef system that isn't stable and is declining as has been shown for other regions such as the Caribbean (Schutte et al 2010).
Sweatman et al: In the kind of broad analysis presented here, reef resilience is manifested as substantial increases in coral cover following disturbance. In the great majority of sub- regions of the GBR, reefs showed both declines and sub- stantial periods of increasing living coral cover over the 19 years of surveys, evidence that many reefs retained their regenerative capacity.This, I largely agree with. In fact, there isn't much doubt that reefs on the GBR have not lost all their regenerative capacity, eg, see here.
Shifting baseline syndrome
In his classic 1995 paper in Trends in Ecology and Evolution, Daniel Pauly outlined his argument for shifting baseline syndrome in fisheries:
Essentially, this syndrome has arisen because each generation of fisheries scientists accepts as a baseline the stock size and species compostion that occurred at the beginning of their careers, and uses this to evaluate changes. When the next generation starts its career, the stocks have further declined, but it is the stocks at that time that serve as a new baseline. The result obviously is a gradual shift of the baseline, a gradual accommodation of the creeping disappearance of resource species, and inappropriate reference points for evaluating economic losses resulting from overfishing, or for identifying targets for rehabilitation measures.The Sweatman et al paper is a great example of shifting baseline syndrome: coral reef scientists that accept as a baseline the state and coral reef cover that occurred at the beginning of their careers and use this to evaluate changes regardless of valid evidence of previously higher estimates.
In conclusion: The science certainly isn't settled and I welcome this and future contributions by Sweatman et al and others on the matter. I started working on this problem about 7 years ago and I frequently ask more experienced scientists and old-timers what they remember from the 60s and 70s and what they think the baseline distribution for coral cover - which certainly varies among regions - really is. We can't know for sure without a time machine. But based on all the data at hand, I feel confident the GBR historically (before people starting mucking it up) had at least twice the coral it has now (including the nearshore reefs that we have lost and don't even bother to survey anymore). This is pretty much what the vast amount of data (10,000+ surveys) for reefs around the world indicates has happened globally. In Australia, this topic matters a lot because there has been an ongoing argument about how much the GBR has changed and how threatened it really is. Sadly, much of this is playing out in the dodgy Aussie newspaper, the Australian. See past debunking of this nonsense about the GBR being "blue again" here, here, here, here and here.
Some background and disclaimers: The lead author, Hugh Sweatman is a colleague and collaborator of mine. We have published several papers together and are working on other collaborative projects and started corresponding about coral reef baselines about five years ago. Dr. Sweatman is the director of the long-term monitoring program at AIMS (the analysis was based on the dataset from this program). I was a reviewer of the manuscript and made the same points in my signed (non-anonymous) review that I made above. Sweatman et al (2011) directly criticized findings of a paper on which I was the lead author (Bruno and Selig 2007). Finally, an expanded version of this post was recently published on ClimateShifts here. - JB
References
Connell J.H. (1978). Diversity in tropical rain forests and coral reefs. Science, 199, 1302-1310
Endean R. (1977). Acanthaster planci infestations of reefs of the Great Barrier Reef. Third International Coral Reef Symposium, 185-191
Endean R. & Stablum W. (1973). The apparent extent of recovery of reefs of Australia's Great Barrier Reef devastated by the crown-of-thorns starfish. Atoll Research Bulletin, 168, 1-41
Done T. (1992). Constancy and change in some Great Barrier Reef coral communities: 1980-1990. American Zoologist, 32, 655-662
Done T.J., Dayton, P.K., Dayton, A.E., Stege, R. (1991). Regional and local variability in recovery of shallow coral communities: Moorea, French Polynesia and central Great Barrier Reef. Coral Reefs, 9, 183-192
Gomez, E. D., A. C. Alcala, and A. C. San Diego. (1982) Status of the Philippine coral reefs – 1981. Proceedings of the Fourth International Coral Reef Symposium, Manila 1:275-282

 0
0  0
0





Comments